Predator-prey Metapopulations
Current work investigates the role of environmental variability in predator-prey metapopulations. In 1953 Moran proposed that the action of widespread meteorological factors could synchronize fluctuations in separate local populations. Metapopulation theory also proposes that regional persistence arises because of some degree of independence in the dynamics of different local populations. Regional metapopulation persistence might therefore be cut short by regional synchronization of population fluctuations. The exact links between environmental variation and predator-prey metapopulation processes are poorly understood, mainly because of the difficulty of studying species interactions and movement in patchy habitats in the field. Alan Hastings has also shown that long-term transient dynamics might become the dominant population processes that we observe in multipatch population systems with nonlinear dynamics within patches (Hastings 2001). This project, in collaboration with Alan Hastings, combines microcosm experiments with modeling and using techniques from phase dynamics and time series analysis to quantify the effects of environmental variation on synchrony (= phase), transient dynamics and regional persistence. The project will both generate new tools to use for quantifying synchrony and answer some unexplored questions about synchrony and metapopulations.
My work on predator-prey metapopulations builds on my earlier research with Sharon Lawler (Entomology, UCD) demonstrating the feasibility of testing metapopulation theory in laboratory microcosms (Holyoak and Lawler 1996a, 1996b). Such theory has proven difficult to test in field systems. An NSF grant supported this work (1996-2000). Recent achievements include demonstrating that the regional persistence of a species can depend on the spatial arrangement of habitat patches if only a few habitat patches are available; this finding is relevant to habitat conservation (Holyoak 2000a). Other investigations showed that predator-prey dynamics changed as the number of dispersal corridors increased, shifting from local populations that experienced semi-independent extinction and colonization, through local populations that were rescued from extinction by immigration, to regionally synchronous and short-lived populations when patches were connected by frequent dispersal (Holyoak 2000a). Such a demonstration of persistence mechanisms is lacking from most spatial dynamics studies. This work is being extended to test ideas about how environmental variation influences spatial dynamics.
I also investigated the effect of spatial dynamics on the 'paradox of enrichment', where improved nutrient conditions for prey paradoxically destabilize predator-prey interactions (Holyoak 2000b). An experiment showed that although nutrient enrichment destabilized local populations, subdivided populations could persist because local extinctions were asynchronous. This might explain why extinctions attributable to the paradox of enrichment are commonly observed in simple models and laboratory experiments, but have been difficult to document in nature.
In collaboration with Sharon Lawler (Entomology, UCD) and Philip H. Crowley (U. of Kentucky), I also investigated how an individual-based model can be used to predict both temporal dynamics and persistence of a predator and prey in single habitat patches (Holyoak, Lawler and Crowley 2000). This helps to improve the testability of persistence mechanisms. Testing persistence mechanisms is difficult in cyclical populations because both intrinsic density changes and stochasticity contribute to extinction. This work, is unusual in that independent experiments parameterized and tested the model.
Another set of experiments, in collaboration with a Ph.D. student A. Amezcua, comprised the first experimental investigation of whether 'source and sink' prey population structure can stabilize predator-prey dynamics (Amezcua and Holyoak 2000). Many species live in patchy habitat where population growth varies from positive to negative ('source' and 'sink' patches). The dynamic consequences of this are largely unknown, aside from theoretical speculation. Our experiments showed that the variation in patch quality could stabilize a predator-prey interaction, in a similar way to the stabilizing effects of asynchronous dynamics in metapopulations. Related experiments (with an ecology Ph.D. student M. Donahue) investigated patterns of between-patch movement and dynamics for a single species exposed to various spatial distributions of resources.
References cited
Amezcua, A. B., and M. Holyoak. 2000. Empirical evidence for predator-prey source-sink dynamics. Ecology 81:3087-3098.
Hastings, A. 2001. Transient dynamics and persistence of ecological systems. Ecology Letters 4:215-220.
Holyoak, M., and S. P. Lawler. 1996a. Persistence of an extinction-prone predator-prey interaction through metapopulation dynamics. Ecology 77:1867-1879.
Holyoak, M., and S. P. Lawler. 1996b. The role of dispersal in predator-prey metapopulation dynamics. Journal of Animal Ecology 65:640-652.
Holyoak, M. 2000a. Habitat patch arrangement and metapopulation persistence of predators and prey. The American Naturalist 156:378-389.
Holyoak, M. 2000b. The effects of nutrient enrichment on predator-prey metapopulation dynamics. Journal of Animal Ecology 69:985-997.
Holyoak, M., S. P. Lawler, and P. H. Crowley. 2000. Predicting extinction: Progress with an individual-based model of protozoan predators and prey. Ecology 81:3312–3329.
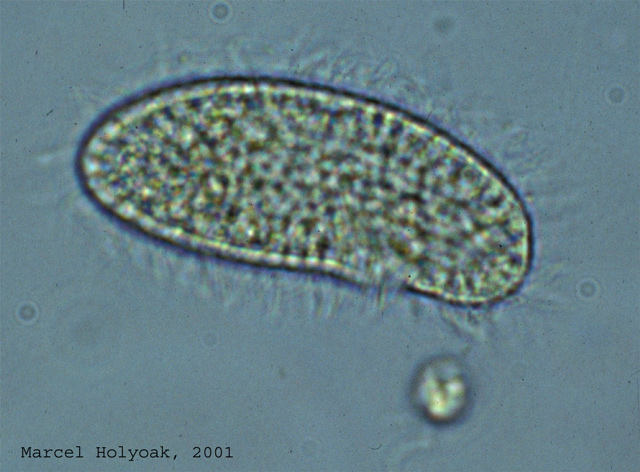
The prey, Colpidium cf.
striatum, a small bacterivorous ciliated protist (length c. 0.05mm).
Click on the picture for a larger image.
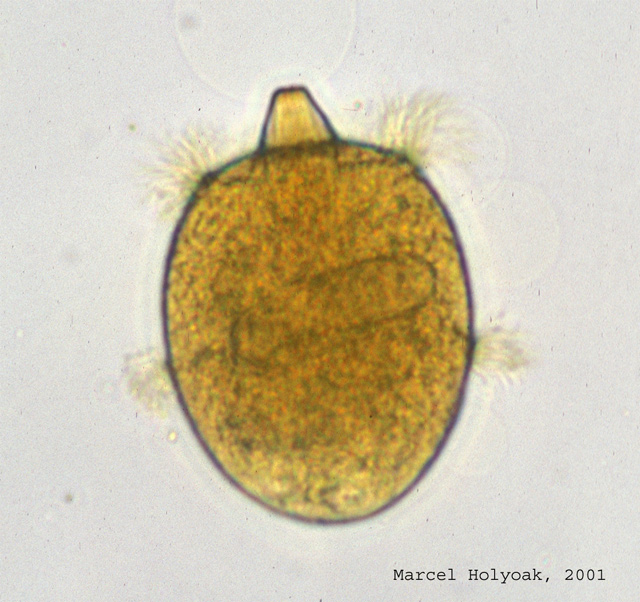
The predator, Didinium nasutum, a medium sized predatory ciliated protist (length c. 0.1mm). Click on the picture for a larger image.

My microcosms typically consist of arrays of interconnected 30-mL bottles, isolated 30-mL bottles, or large undivided bottles of the same total habitat size (not shown). The predators and prey both move freely through the interconnecting tubes.